|
Рост древесного растения обычно начинается с прорастания самого важного органа размножения, семени. Существенная часть семени - живой зародыш, поэтому основное внимание при обработке и хранении семян уделяется созданию условий, обеспечивающих жизнеспособность зародыша, хорошую всхожесть семени.
Возобновление роста зародыша и его развитие в новый, независимый сеянец включают наиболее важные процессы, которые являются предметом изучения физиологии растений. К ним относятся: поглощение воды, усвоение питательных веществ, синтез ферментов и гормонов, азотный и фосфорный метаболизм, передвижение веществ и ассимиляция.
СТРОЕНИЕ И СОСТАВ СЕМЯН
Семена древесных растений различаются размерами, формой, цветом и строением. Размеры бывают от едва видимых, например у оксидендрума древовидного и рододендрона, до таких, как у кокоса, вес которых достигает иногда 9 кг. Поверхность оболочки семян также очень разнообразна: она может быть совершенно гладкой и очень грубой, шероховатой. Семена иногда снабжены дополнительными образованиями, такими, как крылатки, присемянники, выступы, шины, бугорки, волоски.
Нормальное семя - это оплодотворенная зрелая семяпочка, которая содержит зародыш, запасные питательные вещества (лишь очень редко они отсутствуют) и защитную оболочку или оболочки. Термин семя не всегда ограничивается этим определением. Охотнее его используют в функциональном смысле как единицу рассеивания, распространения. В этом смысле термин семя применим к сухим, односемянным или редко дву- многосемянным плодам, также и к настоящим семенам. Например, сухие односемянные плоды (крылатки ильма, орешек и плюска бука) обычно, хотя и неточно, относят к семенам.
Зародыш - это миниатюрное растение, имеющее одну или более семядолей (первых листьев), зародышевые почки, гипокотиль (часть стебля) и корешки (зачаточные корни) (рис. 14.1). Размер зародыша значительно варьирует у семян разных видов. У некоторых видов - это зачаточная структура, у других он почти полностью занимает семя (рис. 14.2). Зародыши семян бамбука и пальмы имеют одну семядолю, поэтому классифицируются как однодольные. Зародыши большинства древесных покрытосеменных имеют две семядоли, у голосеменных, в зависимости от вида, возможны от 2 до 18 семядолей.
 Зрелое семя сосны Ламберта (Anonymous, 1948) :
а-б - внешний вид в двух планах; в - продольный разрез; г - зародыш: 1 - кожура семени; 2 - нуцеллус; 3 - почечка зародыша; 4 - семядоли; 5 - эндосперм; 6 - полость зародыша; 7 - первичный корешок; 8 - подвесок; 9 - микропиле Зрелое семя сосны Ламберта (Anonymous, 1948) :
а-б - внешний вид в двух планах; в - продольный разрез; г - зародыш: 1 - кожура семени; 2 - нуцеллус; 3 - почечка зародыша; 4 - семядоли; 5 - эндосперм; 6 - полость зародыша; 7 - первичный корешок; 8 - подвесок; 9 - микропиле
 Строение семени (Anonymous, 1948):
а - аралии, большой эндосперм и маленький зародыш; б - тсуги, большой зародыш, окруженный эндоспермом; в - ирги канадской, эндосперма нет, зародыш почти целиком заполняет полость семени; 1 - наружная кожура семени; 2 -внутренняя кожура семени; 3 - эндосперм; 4 - семядоля; 5 - первичный корешок ; 6 - микропиле Строение семени (Anonymous, 1948):
а - аралии, большой эндосперм и маленький зародыш; б - тсуги, большой зародыш, окруженный эндоспермом; в - ирги канадской, эндосперма нет, зародыш почти целиком заполняет полость семени; 1 - наружная кожура семени; 2 -внутренняя кожура семени; 3 - эндосперм; 4 - семядоля; 5 - первичный корешок ; 6 - микропиле
Питательные вещества в семенах могут быть локализованы в семядолях или в тканях, окружающих зародыш, у покрытосеменных - в эндосперме. Нормальный эндосперм по числу хромосом тригатоиден, так как образуется после соединения диплоидного ядра и спермия. Эндосперм - основная запасающая питательные вещества ткань семян многих двудольных видов. У семян некоторых растений, таких, как айлант, некоторые питательные вещества запасаются в эндосперме,
другие - в семядолях. В семенах голосеменных питательные вещества запасаются прежде всего в мегагаметофите (женском гаметофите), который окружает зародыш. Мегагаметофит по числу хромосом гаплоиден и отличается по происхождению от эндосперма, хотя и выполняет те же функции.
Семенная кожура, защищающая зародыш от высыхания или от поражения вредителями, состоит из наружной твердой оболочки, тесты и тонкой внутренней пленчатой оболочки. Имеются значительные различия в свойствах кожуры семян. Например, у тополя и ивы теста очень мягкая, а у боярышника, падуба и большинства бобовых она очень твердая. У ильма и внутренняя и наружная оболочки семян пленчатые. Простая кожура семян голосеменных может быть твердой, как у сосны, или мягкой, как у пихты.
ПРОДОЛЖИТЕЛЬНОСТЬ ЖИЗНИ СЕМЯН
Одна из самых характерных особенностей семян - широкая вариабельность продолжительности жизни, которая может быть от нескольких дней до нескольких десятилетий и даже столетий. Известно, например, что семена люпина из арктической тундры, которые были погребены в замороженном иле на протяжении 10 тыс. лет, хорошо проросли в лаборатории. Семена таких древесных пород, как тис, тополь, ильм, ива, дуб, гикори, береза и конский каштан, содержат много воды и недолговечны. Быстро теряют всхожесть семена многих тропических растений. К ним относятся представители таких родов: Theobroma, Coffea, Cinchona, Erythroxylon, Litchia, Monterzuma, Macadamia, Hevea, Thea и Cocos. Однако подбором подходящих температуры и влажности во время хранения семян можно продлить срок их жизни от нескольких недель или месяцев до года.
Семена постепенно стареют и переходят от одной стадии нарушения жизнедеятельности к другой. Начальные симптомы старения семян заключаются в неспособности прорастать и увеличении чувствительности к микроорганизмам. По мере ухудшения состояния семян с возрастом у них при прорастании появляются короткие корешки, а семядоли не могут пробиться через оболочку семян. В конце концов семена погибают. Скорость старения семян контролируется в основном влажностью среды, от которой зависит содержание воды в семени, и температурой, которая влияет на биохимические процессы. Гарингтон (1972) ссылается на два полезных правила, оба убедительны и применяются независимо друг от друга. Одно из них касается влияния на старение семян влажности в сухих семенах, другое - температуры:
- При увеличении содержания воды в семенах на 1% (в диапазоне от 5 до 14% исходной их влажности) срок жизни семян сокращается в два раза. При содержании воды в семенах ниже 5% старение часто ускоряется из-за самоокисления липидов. При содержании воды в семенах выше 14% прорастание семян часто снижается из-за поражения их грибами.
-
При увеличении температуры на каждые 5°С срок жизни семян уменьшается в два раза. Это правило обычно применимо при повышении температуры от 0 до 50°С.
ТИПЫ ПРОРАСТАНИЯ СЕМЯН
Прорастание семян можно рассматривать как начало роста зародыша, в результате чего происходят разрыв оболочки семени и появление молодого растения. Рост зародыша требует и деления клеток и растяжения, причем у одних видов сначала происходит деление клеток, у других вначале наступает растяжение клеток. Например, деление клеток предшествует их растяжению во время роста зародыша семян Pinus thunbergii. У лавровишни, однако, в эмбриональных органах деление и растяжение клеток начинаются почти одновременно. Запасные питательные вещества в семени поддерживают растущий зародыш до тех пор, пока развернутся листья - фотосинтезирующая система - и разовьются корни, способные поглощать воду и минеральные вещества. Таким образом, молодое растение становится физиологически самостоятельным.
В процессе прорастания семени корешок удлиняется и проникает в почву. У некоторых древесных растений, таких, как большая часть видов голосеменных, бук, кизил, белая акация, ясень и многие виды клена, семядоли пробиваются наверх при помощи растущего гипокотиля (надземное прорастание). У других видов, таких, как дуб, орех, конский каштан, семядоли остаются под землей, в то время как эпикоптиль растет вверх и развивает листья (подземное прорастание).
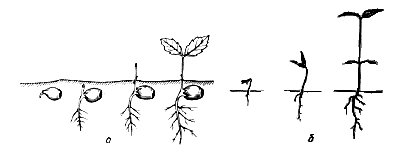 Стадии прорастания семян (переснято из разных источников) :
а - желудя дуба белого, у которого семядоли остаются под землей (подземное)
б - клена красного, у которого семядоля пробиваются на поверхность почв
(надземное) Стадии прорастания семян (переснято из разных источников) :
а - желудя дуба белого, у которого семядоли остаются под землей (подземное)
б - клена красного, у которого семядоля пробиваются на поверхность почв
(надземное)
 Надземное прорастание семян сосны (фото t-Regis Paper Co.)
Надземное прорастание семян сосны (фото t-Regis Paper Co.)
ВЛИЯНИЕ ОКРУЖАЮЩИХ УСЛОВИЙ НА ПРОРАСТАНИЕ СЕМЯН
Быстрое прорастание семян очень желательно, так как при этом уменьшается угроза повреждения семян насекомыми, грибами или неблагоприятными условиями, а также поедания их птицами или грызунами. Из окружающих факторов, контролирующих прорастание семян, очень важными являются вода, температура, свет, кислород и различные химические вещества.
ВОДА. Семена, выходя из состояния покоя, должны поглотить некоторое количество воды для того, чтобы у них возобновились физиологические процессы, связанные с прорастанием. Например, дыхание семян значительно возрастает при небольшом возрастании гидратации выше критического уровня. Абсолютное количество воды, необходимое для начала прорастания, относительно невелико, как правило, не превышает 2-3-кратного веса семян. Растущий проросток нуждается в большом количестве воды, и эта потребность возрастает с усилением транспирации. Все жизнеспособные семена, за исключением тех, которые обладают непроницаемой оболочкой, при прорастании могут поглощать воду из почвы при полевой влагоемкости. На подсыхающих почвах скорость прорастания и конечный процент проросших семян снижается.
Влияние влажности почвы на прорастание семян зависит от их принадлежности к определенным видам. Когда влажность почвы становилась ниже полевой влагоемкости, прорастание семян сосны густоцветной и кипарисовика туполистного снижалось. При этом семена кипарисовика были более чувствительны к подсушиванию. Например, всхожесть семян кипарисовика туполистного понижалась примерно на 6% на каждый бар снижения водного потенциала почвы. В этих условиях у сосны густоцветной всхожесть семян падала лишь на 2,5%. Влияние водного стресса на прорастание семян часто зависит от температуры. Например, при температуре около 38°С водный стресс был решающим фактором в самом начале прорастания семян мескитового дерева. Однако при 29°С на рост зародыша сильнее влияла температура, а водный стресс становился лимитирующим фактором только в конце процесса прорастания.
ТЕМПЕРАТУРА. Из состояния покоя семена выводятся низкими температурами, а для индуцирования быстрого прорастания необходимы значительно более высокие температуры. Семена с нарушенным покоем могут расти и при низкой температуре, но для начала ростовых процессов требуется больше времени. Прорастание любой партии семян может происходить в широком температурном диапазоне, но в его пределах имеется оптимум, при котором в кратчайший срок достигается самый высокий процент всхожести. Минимальная, оптимальная и максимальная температуры прорастания семян значительно различаются у разных видов. У видов умеренной зоны они обычно ниже, чем у тропических. Температуры прорастания также значительно зависят от места сбора семян. Оптимальные температуры бывают различными и для семян, собранных с разных деревьев одного и того же вида.
Семена многих видов часто прорастают одинаково хорошо в широких пределах температурной шкалы. Семена сосны скрученной широкохвойной прорастали примерно с одинаковой скоростью при 20° и 30°С, у семян сосны Банкса не отмечалось заметных различий в скорости прорастания при 15, 21 и 27°С при непрерывном освещении. Есть данные о том, что массовое появление проростков ели Энгельмана и сосны скрученной широкохвойной было и при 16°, и при 25°С, но меньше всего при 35°C. Однако при 16°С проростки ели появились на два дня раньше, чем проростки сосны. У некоторых видов семена прорастают в узком температурном диапазоне, который можно изменить только применением предпосевной обработки семян. Например, семена клена платановидного прорастали лучше всего при 5° и 10°С. Всхожесть семян клена ясенелистного составляла 67% при температурах от 10 до 25°С, и лишь 12% при температуре между 20 и 25°C.
Семена многих видов хорошо прорастают при постоянной температуре, но обычно все-таки необходимы суточные температурные колебания. Так, семена ясеня маньчжурского увлажняли и выдерживали при низкой температуре, а затем при 25°С. При этом некоторое количество семян прорастало, а при постоянной температуре 8°С прорастание существенно тормозилось. При содержании семян в течение 20 ч при 8°С и затем при температуре до 25°С на протяжении 4 ч прорастание их значительно ускорялось. Прорастание семян сосны густоцветной также ускорялось под влиянием суточных колебаний температур.
РАДИАЦИЯ. Большинство семян не чувствительно к свету и прорастает одинаково хорошо как в темноте, так и на свету. Семена некоторых видов требуют для прорастания свет. Так, для ели необходимо освещение всего в 0,08 лк, для березы 1 лк, для сосны 5 лк. Но есть и такие виды, для прорастания семян которых необходимо освещение до 100 лк. Интенсивность освещения оказывает относительно небольшое влияние на прорастание, но часто длина дня и длина волны усиливают этот эффект. Стимулирующее действие света на прорастание обусловлено увеличением потенциала ростовых процессов зародыша.
Длина дня. Для массового прорастания семян большинства светочувствительных видов древесных растений необходим 8-12-часовой фотопериод. Прерывание темнового периода короткими световыми вспышками, а также увеличение температуры обычно оказывают то же действие, что и удлинение экспозиции на свету. Можно привести несколько примеров различных требований к фотопериоду при прорастании семян разных видов. У тсуги восточной 8 или 12-часовой день вызывал максимальное прорастание семян, с увеличением дня до 14 или 20 ч дополнительный эффект не наблюдался. Семена эвкалипта хорошо прорастали при 8-часовом дне, семена березы - при 20-часовом фотопериоде. Семена дугласии, однако, прорастали на длинном, или 16-часовом, дне, но совсем не прорастали при 8-часовом фотопериоде.
Длина волны. Прорастающие семена большого числа видов травянистых и древесных растений чувствительны к свету определенной длины волны. Это характерно для таких древесных покрытосеменных, как Alnus inocuma, Artemisia monosperma, Betula pubescens, Fraximus mandshunca и Ulmus americana. Среди голосеменных к свету определенной длины волны при прорастании семян чувствительны Abies, Picea и Pinus (P. thunbergii, P. strobus, P. palustris, P. silvestris, P. taeda и Р. virginiana).
Ближний красный свет стимулирует прорастание дальний ингибирует его. Если ближний (650 нм) и дальний красный (730 нм) свет дают последовательно, то способность семян прорастать зависит от последней длины волны. Стимулирующее влияние красного света зависит часто от температуры или длительности снабжения водой. Например, установлено, что прорастание стимулированных красным светом семян сосны виргинской ускорялось больше после 20-дневного намачивания в воде при 25°С, чем после 1-дневного намачивания. Прорастание семян стимулировалось красным светом сильнее, если перед облучением они поглощали воду при 5°С, а не при 25°С.
КИСЛОРОД. Дыхание необходимо на ранней фазе прорастания семян, поэтому снабжение кислородом влияет на прорастание. Семена обычно нуждаются в более высоких концентрациях кислорода для прорастания, чем это требуется для последующего роста проростков. Относительно высокая потребность в кислороде у семян некоторых видов растений объясняется наличием кожуры, являющейся барьером для диффузии кислорода в семена. Удаление кожуры семян сосны смолистой или выдерживание интактных семян при высокой концентрации кислорода значительно увеличивало скорость поглощения кислорода. Выдерживание семян, освобожденных от кожуры, при высокой концентрации кислорода еще больше ускоряло дыхание.
Замачивание семян в течение нескольких часов ускоряет прорастание, но продолжительное замачивание индуцирует повреждение и потерю жизнеспособности многих семян, главным образом вследствие сниженной концентрации и доступности растворенного кислорода. Степень влияния замачивания семян на прорастание значительно отличается у разных видов растений. Замачивание семян некоторых видов в течение 3-5 дней не снижало всхожести, но замачивание в течение 10 дней значительно ухудшало прорастание, удлинение срока ,до 30 дней убивало семена. Семена растений, произрастающих в долинах (например ниссы и болотного кипариса), обладали малой потребностью в кислороде и могли выдерживать продолжительное погружение в воду, не теряя всхожести. Не обнаружено существенного влияния на прорастание у 6 видов растений долин после замачивания семян в течение 32 дней.
СРЕДЫ ДЛЯ ПОСЕВА СЕМЯН. Прорастание семян и укрепление проростков в естественных условиях в значительной мере зависят от их физических свойств, температуры, доступности воды и минеральных веществ. Почва является хорошей средой для посева семян благодаря высокой способности к фильтрации воды, хорошей аэрации и тесному контакту между частицами почвы и семенами. Остатки растений и лесная подстилка менее пригодны для посева, так как они медленно прогреваются, препятствуют проникновению корней и контакту семян с минеральными веществами почвы, быстро высыхают и затеняют маленькие проростки. Сфагнум, обладающий высокой водоудерживающей способностью, часто пригоден в качестве среды для посева, но после их прорастания он может заглушить молодые растения. Разлагающаяся древесина также прекрасный естественный субстрат для прорастания семян лесных деревьев из-за ее способности поглощать и удерживать влагу.
ФИЗИОЛОГИЯ ПРОРАСТАНИЯ СЕМЯН
Основной момент в прорастающем семени - начало роста зародыша и его превращение в самостоятельное растение. Когда начинается прорастание, включаются многие процессы, при этом происходит:
- гидратация семени;
- увеличение скорости дыхания;
- активация ферментов;
- увеличение количества аденозинфосфатов;
- увеличение количества нуклеиновых кислот;
- усвоение запасных питательных веществ и транспорт растворимых продуктов к зародышу, где синтезируются клеточные компоненты;
- активация деления клеток и их растяжение;
- дифференциация клеток в ткани и органы.
Точный порядок ранних изменений не ясен, наблюдаются наложения одного этапа на другой, но первым обязательным этапом всегда является поглощение воды. Увеличение гидратации связано с растяжением и делением клеток в точках роста, так же как и с освобождением гормонов, которые стимулируют образование ферментов и их активность. При набухании увеличивается сырой вес семени и одновременно вследствие окисления субстратов и некоторой утечке их из семян снижается сухой вес. Когда появляются корни и начинают поглощать минеральные вещества, а семядоли или листья становятся фотосинтетически активными, сухой вес проростков вновь увеличивается и достигает исходного уровня, а затем даже превосходит первоначальный вес семян.
ПОКОЙ СЕМЯН
Зрелые семена многих древесных растений, попав в соответствующие условия, сразу же прорастают, но семена большинства видов проходят через стадию покоя. Это означает, что они не прорастают сразу после созревания даже если их посеять в самые благоприятные условия. Покой семян может быть для растений благоприятен и неблагоприятен. Для прохождения стадии покоя семенам многих растений умеренной зоны требуется продолжительное охлаждение, поэтому mm не прорастают до наступления весны. Это обеспечивает молодым растениям безопасность, так как раннее прорастание привело бы проростки к гибели от морозов. Семена некоторых дикорастущих растений могут оставаться в состоянии покоя в почве на протяжении многих лет, и период прорастания таких семян растягивается на годы. Такой длительный покой способствует закреплению и выживанию видов, даже если случайно очень рано появившиеся некоторые проростки данного года будут убиты засухой или морозом. Другое преимущество покоя семян в жарких и сухих областях заключается в том, что они прорастают в очень короткий влажный период года. У семян некоторых пустынных растений ингибиторы, содержащиеся в оболочке, препятствуют прорастанию. Однако ингибиторы выщелачиваются обильными дождями во влажную почву, вследствие чего семена прорастают лишь при достаточном для укоренения растений содержании воды в почве. С другой стороны, работники питомников рассматривают покой семян как отрицательное явление, так как они стремятся быстро получать семена хорошей всхожести, чтобы иметь большой и однородный выход проростков. Причины покоя семян и методы нарушения его имеют физиологическое и практическое значение.
ПРИЧИНЫ ПОКОЯ. Знание причин покоя семян часто способствует целесообразному применению соответствующих приемов для преодоления состояния покоя отдельных партий семян. Покой семян обусловлен рядом причин:
- незрелостью зародыша;
- непроницаемостью оболочек семени;
- механическим сопротивлением оболочек семени растущему зародышу;
- блокированием метаболических процессов в зародыше;
- комбинацией первого и второго условий;
- вторичным покоем.
Семенная кожура и покой семян. Распространенная причина покоя - непроницаемость оболочек для воды и кислорода. Покой семян, связанный со свойствами оболочек, особенно характерен для семян растений семейства Leguminosae, таких, как белая акация, гледичия и церцис. Проблема оболочек семян также характерна для можжевельника виргинского, липы, сосны веймутовой и яблони. Природа непроницаемости семенных оболочек у разных видов различна. У гледичии и белой акации для всасывания воды и поглощения кислорода семенная кожура является барьером. У сосны веймутовой и ясеня зеленого кожура проницаема для воды, но препятствуют газообмену. Кожура семян яблони задерживает поглощение кислорода, создавая его недостаточность, что препятствует высокой скорости дыхания, необходимой при прорастании зародыша.
Считается, что семенная кожура некоторых видов механически препятствует прорастанию зародыша, и он полностью развивается внутри семени. Однако Виллерс (1972) утверждает, что причины этого явления во многих опубликованных случаях иные, возможно, это обусловлено физиологическим покоем зародыша.
Покой зародыша. Иногда зародыш не созревает до конца вегетационного периода и требует времени для дозревания (хранения при благоприятных условиях), чтобы достичь определенной стадии развития и быть готовым к прорастанию (например, семена калины, падуба и гинкго). Однако наиболее распространен такой тип покоя семян, при котором морфологически зрелый зародыш не может возобновить рост и прорастать. Этот ршп физиологического покоя зародыша распространен среди покрытосеменных у видов яблони, сирени, дуба, каштана, кизила, гикори, груши и явора. У голосеменных физиологический покой зародыша встречается у некоторых видов сосны, кипариса болотного, дугласии, тсуги, можжевельника, лиственницы, ели и пихты.
Состояние физиологического покоя зародыша, так же как и спящих почек, проходит, по-видимому, в две стадии, одна из которых - слабый и обратимый покой - постепенно переходит в глубокий покой, который необратим в тех же условиях, которые его вызвали. Семена некоторых видов Fraxinus, нормально переходящие в состояние покоя, начинают прорастать сразу же, если их собрать и посеять до того, как они успевают пройти через фазу высушивания. Иногда неспособность семян прорастать обусловлена воздействием нескольких типов покоя. У некоторых видов Rosa прорастанию препятствуют как механическое сопротивление мощного перикарпия увеличивающемуся в размерах зародышу, так и покой семян, который возникает под влиянием ингибиторов роста. Такой "удвоенный покой" известен также для семян боярышника, можжевельника виргинского, тиса, липы, дерена, маклюры яблоконосной, скумпии американской, гамамелиса виргинского, сосен Сабина, кедровой европейской и белоствольной. Покой семян падуба обусловлен как незрелостью зародыша, так и твердой кожурой семян.
Гормоны и покой семян. Весь физиологический покой семян, вероятно, находится под контролем ростовых гормонов. Начало покоя часто связывают с накоплением ингибиторов, а нарушение покоя - со сдвигом в балансе гормонов в пользу стимуляторов роста, которые начинают перекрывать действие ингибиторов. Сходство состояния покоя зародышей семян и покоя почек подчеркивается тем, что физиологические карликовые проростки, образующиеся из неохлажденных зародышей, очень похожи на ветви плодовых деревьев с розеточной формой роста после мягкой зимы. Кроме того, и покой зародышей и покой почек могут быть нарушены охлаждением, экзогенными стимуляторами роста и длинным фотопериодом.
Стимуляторы роста. Важность стимуляторов роста в преодолении покоя семян может быть иллюстрирована несколькими примерами. Как уже ранее упоминалось, в процессе нормального дозревания или после искусственного охлаждения некоторых семян уровень стимуляторов роста возрастает, хотя количество ингибиторов не всегда снижается. Выщелачивание семян, которое, как предполагается, стимулирует прорастание вследствие вымывания ингибиторов, часто вызывает формирование карликовых проростков, вероятно, за счет удаления и стимуляторов роста. Например, примененный гиббереллин индуцировал ферментативную активность и рост зародыша у лещины. У клена белого цитокинины, а не гиббереллины, нарушали покой семян. У других видов, таких, как персик и клен сахарный, гиббереллины и цитокинины могли преодолеть покой семян. Эти эксперименты подчеркивают сложность и изменчивость специфического гормонального регулирования покоя зародышей у разных видов. Кан (1975) подчеркивает возможность гормональных взаимоотношений в покоящихся семенах и предлагает приемлемую модель, в которой гиббереллины имеют первостепенное значение в переходе семян к прорастанию, а роль ингибиторов и цитокининов рассматривается как вторичная, так как их действие вполне предотвратимо.
НАРУШЕНИЕ ПОКОЯ. У многих видов покой семян может быть нарушен приемами, направленными на изменение соотношения между ингибиторами и стимуляторами роста и увеличивающими их механическое сопротивление по отношению к растущему зародышу. Эффективность применяемого воздействия зависит от глубины и характера покоя семян. У одних видов покой семян легко нарушить любой обработкой, семена других видов отзываются лишь на какой-нибудь один, специальный прием. Но есть и такие виды, семена которых не удается вывести из состояния покоя ни одним из известных и широко применяемых методов.
Приемы, используемые для нарушения покоя зародыша, обычно также направлены на то, чтобы сдвинуть соотношение между гормонами в пользу стимуляторов роста. Это может сопровождаться либо снижением уровня эндогенных ингибиторов, либо увеличением уровня стимуляторов роста. Так, по исследованиям ряда авторов, охлаждение семян персика и ясеня резко снижало уровень ингибитора (АБК). Однако снижение уровня ингибиторов не всегда объясняет прекращение покоя семян, которые были обработаны охлаждением. Брадбером (1958) показано, что воздействие низкими температурами нарушает покой семян лещины вследствие синтеза гиббереллина.
Дозревание. У семян, обладающих лишь средней глубиной покоя зародыша, период дозревания проходит в условиях сухого хранения. Во время дозревания устойчивость семян к окружающим условиям увеличивается. Температурный диапазон для, прорастания становится значительно шире по сравнению с температурами для свежесобранных семян, которые могут прорастать либо при очень низких, либо при очень высоких температурах. Специфические требования к температурному и световому режиму в процессе дозревания постепенно исчезают.
Скорость дозревания зависит от времени года и вида растений. Даже у генетически однородной популяции семян, которые дозревают в одних и тех же окружающих условиях, отдельные семена бывают разной степени зрелости. У некоторых видов семян при сухом хранении происходит множество различных физических и химических изменений. По крайней мере часть из них является результатом изменений в кожуре семени, которая изменяет свою силу натяжения или увеличивает проницаемость для воды и газов. В других случаях изменения происходят в зародыше или в окружающих тканях. У семян с незрелыми зародышами происходят дальнейшие морфологические преобразования. У некоторых видов ясеня морфологически сформировавшийся зародыш, прежде чем начать прорастать, должен увеличиться в размерах. Биохимические изменения в процессе дозревания включают: уменьшение количества запасных липидов, углеводов и белков; возрастание метаболической активности; изменение гормонального баланса таким образом, чтобы стимуляторы роста преобладали над ингибиторами.
Стратификация. Покой семян обычно нарушается после хранения их в течение 30-120 дней при температурах 1-5°С, при обильной аэрации и влажности. Этот прием воспроизводит до некоторой степени естественные зимние условия. Термин стратификация употребляют в случаях, когда семена помещают между слоями материалов, хорошо удерживающих влагу (сфагнум, опилки, песок). Затем создают низкую температуру. Низкая температура и влажность - необходимые условия для хранения семян слоями. Таким образом, при стратификации семена увлажняют, помещают в пластиковые мешки и хранят при низкой температуре. После такой обработки их можно высевать в землю, но если после этого семена подсушить, то они могут перейти в состояние очень глубокого вторичного покоя.
Химические вещества. Покой зародышей семян часто нарушается такими химическими веществами, как гибберелловая кислота и цитокинины. У видов с относительно слабым покоем зародыша (сосен ладанной, ежовой и густоцветной, лиственниц субальпийской и западной) окислительные вещества, например перекись водорода, стимулировали дыхание и ускоряли прорастание. Однако перекись водорода имеет практические ограничения при стимуляции прорастания некоторых семян с покоящимся зародышем. Например, было показано, что при обработке семян веймутовой сосны концентрированной перекисью водорода у одних партий семян удавалось стимулировать прорастание, а другие семена погибали, вследствие этого конечный процент всхожести этих партий семян снижался.
Скарификация. Покой семян с непроницаемой оболочкой может быть нарушен намачиванием в концентрированной серной кислоте в течение 15-60 мин. Проницаемость кожуры семян можно увеличить прокалыванием или обработкой ее наждаком.
ФИЗИОЛОГИЯ МОЛОДЫХ ПРОРОСТКОВ
Как у голосеменных, так и у покрытосеменных растений семядоли играют первостепенную физиологическую роль в ростовых процессах проростков. Семядоли, которые остаются в земле (у ореха, дуба), служат прежде всего запасающими органами. Наземные семядоли некоторых древесных растений также запасают значительные количества углеводов. У других видов, например у сосны и дерена, в семядолях накапливаются лишь небольшие количества углеводов, но они становятся фотосинтетически активными вскоре после появления на поверхности почвы. У видов, семена которых лишены эндосперма (безбелковые семена), семядоли приспособлены и для запасания и для фотосинтеза (например, у березы и белой акации). Если в семенах имеется эндосперм, то семядоли служат не только как запасающие органы, они поглощают запасные питательные вещества из эндосперма и переносят их к растущим осевым органам.
В семядолях откладываются в разных количествах в запас также и минеральные вещества. Зародышевые семядоли безбелковых семян клена красного и белой акации содержат огромные количества питательных элементов, которые перемещаются к быстро развивающимся осевым органам в течение ранних стадий роста проростков. Напротив, в небольших, тонких семядолях ясеня зеленого откладываются в запас незначительные количества питательных веществ.
Развивающийся проросток - это также сложная система с участками синтеза углеводов, которые в онтогенезе сеянца перемещаются от семядолей к первичной, а затем к вторичной хвое. Существует тесная зависимость роста позднее развивающихся листьев от способности к синтезу необходимых для роста веществ ранее появившихся листовых зачатков. Отсюда развитие первичной хвои зависит от вклада семядолей, а развитие вторичной хвои, в свою очередь, зависит от вклада, внесенного первичной хвоей. Молодой проросток на стадии семядолей функционирует с максимальной потребностью в питательных веществах для роста, поэтому он очень чувствителен к действию факторов среды. В начале онтогенеза окружающие условия сильно влияют на заложение всех органов, кроме ранее сформированных зачатков первичной хвои, и на рост всей первичной хвои, включая и сформированные ранее.
У покрытосеменных рост и развитие проростков на стадии семядолей также необычайно чувствительны к стрессовым условиям, так как они влияют на синтез веществ, необходимых для роста. Большая часть углеводов, используемых в процессе роста, у некоторых древесных покрытосеменных с выходящими на поверхность почвы семядолями поступает именно из семядолей до тех пор, пока не разовьются листья. Маршал и Козловский (1974) показали, что продукты фотосинтеза из семядолей экспортировались к растущим тканям листьев. Лишь полностью развернувшиеся листья лиственных пород приобретали большее значение, чем семядоли в синтезе питательных веществ, которые транспортировались из них ко всем растущим органам. Удаление или притенение семядолей в одинаковой мере ингибировало рост проростков, следовательно, в развитии проростка важную роль играет фотосинтез семядолей.
Начало стадии старения характеризуется прогрессивным пожелтением семядолей, которое продолжается до их опадения. Функциональный жизненный цикл семядолей у разных видов различен. Например, по некоторым данным, надземные семядоли белой акации, шелковицы и черной ивы жили недолго (около одного месяца). Более долговечными были семядоли тополя, хмелеграба и явора (около двух месяцев). Еще дольше жили семядоли аралии колючей, каркаса западного и граба (около трёх месяцев) и необычайно долго жили семядоли ясеня американского и магнолии (более 6 месяцев). Точно установить продолжительность жизненного цикла семядолей у различных видов трудно, так как он в значительной мере зависит от стрессовых условий. Например, засуха может индуцировать быстрое старение и опадение в зрелых семядолей.
24 января 2003 г.
|